

Sommaire
[ad_1]
Les oiseaux utilisent différents types de vocalisations produites dans différents contextes : maintien du contact avec les congénères, alarme en présence d’un prédateur, défense du territoire, attraction d’un partenaire sexuel…
À l’instar de l’espèce humaine, plus de la moitié des espèces d’oiseaux sont capables d’imiter les sons produits par des congénères. Chez de nombreuses espèces, cette capacité dépasse même le cadre du répertoire vocal spécifique : il peut s’agir de sons produits par d’autres espèces, de sons de l’environnement, voire de mélodies musicales.
Jusqu’à quel point les vocalisations produites par les oiseaux peuvent-elles nous renseigner sur leurs capacités cognitives et leur état interne : à quoi pensent-ils ? que ressentent-ils ?
En 1974, le philosophe Thomas Nagel publie un article au titre suivant : « Quel effet cela fait-il d’être une chauve-souris ? ». Selon lui, comme Jakob Von Uexküll qui l’a précédé dans cette réflexion avec la notion d’Umwelt (le monde propre de chaque espèce), nous n’avons aucun moyen de se mettre à la place d’un animal d’une autre espèce et de pouvoir ainsi appréhender son expérience de vie. Mais l’éthologiste possède néanmoins des outils pour étudier la communication chez les animaux et proposer des hypothèses vérifiables par l’expérimentation, à la fois en laboratoire mais aussi sur le terrain.
Avant de s’intéresser aux messages véhiculés par les vocalisations chez les oiseaux, il faut rappeler qu’il en existe près de 10 000 espèces différentes. Chaque espèce représente une histoire différente, bâtie au fil de centaines de milliers voire de millions d’années. Il faut donc prendre en considération les conditions socio-écologiques de chaque espèce et son histoire évolutive pour mieux appréhender ce que ses représentants communiquent vocalement.

Nouveau Larousse illustré, Claude Augé, planche oiseaux (1898)
Talents d’imitation
Comme les humains, près de la moitié des espèces d’oiseaux sont capables d’imiter les sons de leur environnement social, principalement ceux produits par des individus conspécifiques. Ces espèces appartiennent à trois groupes taxonomiques différents : les psittacidés (perruches et perroquets), les trochilidés (colibris) et les oscines (environ les deux tiers des espèces de passereaux tels que le merle noir, le rossignol philomèle, le canari).
Parmi les vocalisations produites par les oiseaux par apprentissage, c’est le chant des oscines qui a fait l’objet du plus grand nombre d’études. Chez le diamant mandarin, on a découvert que seuls les mâles chantent et que le chant est appris pendant une période sensible de la vie précoce, comprise entre 25 et 90 jours environ après l’éclosion.
Chez d’autres espèces comme le canari, le chant peut changer au cours de la vie, d’une saison à une autre. Enfin, si pour certaines espèces, ces changements peuvent s’étaler sur plusieurs semaines voire sur plusieurs mois, certaines espèces sont capables de modifier quasi-instantanément leurs vocalisations pour imiter un modèle. Certaines espèces sont également capables d’imiter des sons de leur environnement sonore. L’apprentissage de mots humains par certaines espèces en est l’exemple le plus emblématique. C’est le cas chez de nombreuses espèces de psittacidés et de corvidés mais aussi chez certaines espèces d’oscines comme l’étourneau sansonnet.
Au Siècle des Lumières, dans les milieux aristocratiques en France et dans d’autres pays européens, on entraînait ainsi des oscines (canaris, rossignols…) à imiter des mélodies musicales.
Décoder les vocalisations
Si des vocalisations ont été décrites chez toutes les espèces d’oiseaux recensées, certaines espèces ne vocalisent pas ou peu alors que d’autres ont un répertoire vocal riche de plusieurs types différents de vocalisations émises dans des contextes différents.
Cette description se trouve facilitée depuis la deuxième moitié du XXe siècle par la possibilité d’enregistrer et d’analyser les productions vocales des oiseaux, voire de les synthétiser avant de les diffuser. Cette procédure permet de tester la réaction des oiseaux à des stimuli naturels ou artificiels, et d’essayer ainsi de décoder le message véhiculé en identifiant les sons ou les paramètres acoustiques pertinents dans la transmission du message.
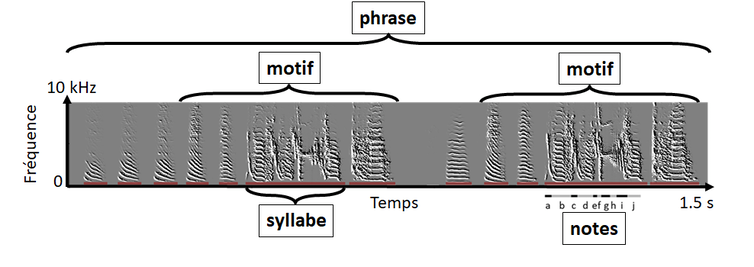
Author provided
Le travail consiste dans un premier temps à décrire le répertoire vocal : classer les vocalisations en fonction de leur contexte d’émission et de leur sémantique, en utilisant parfois des onomatopées et/ou des mots humains pour faciliter leur identification.

Jean‑Pierre Dalbéra/Flickr
En 1871, dans l’ouvrage qu’il consacre à l’évolution humaine et à la sélection sexuelle, Charles Darwin souligne déjà que « les sons émis par les oiseaux offrent, à plusieurs égards, l’analogie la plus proche du langage […] ». Comme pour le langage humain, les vocalisations produites par les oiseaux ont différentes fonctions qui ne sont pas mutuellement exclusives. Parmi les fonctions du langage définies par le linguiste Roman Jakobson en 1963, seule la fonction métalinguistique (la possibilité de parler du code lui-même à savoir le langage chez l’humain) n’est pas observée dans les autres communications animales. La fonction expressive fait référence aux motivations de l’émetteur et dépend aussi de ses caractéristiques intrinsèques, de ce que l’on résume comme l’état interne de l’animal : son état physiologique, ses émotions.
L’influence des états émotionnels
Si certaines vocalisations peuvent être émises dans des situations différentes (par exemple des cris de contact lors de la communication entre individus d’un groupe social, ou des cris d’alarme lors de la détection d’un prédateur), la structure acoustique de certains cris peut être modifiée en fonction de l’état émotif de l’émetteur. Par exemple, chez le diamant mandarin mâle, la structure spectrale du cri de contact produit en présence d’un congénère est différente de celle du cri produit lorsque l’émetteur est isolé et stressé. La diffusion de ces cris de contact produits par un individu stressé entraîne une augmentation de corticostérone, l’hormone du stress, chez les oiseaux testés, ce que l’on peut caractériser comme une contagion émotionnelle.
Toujours dans le registre des émotions, des développements récents en neurobiologie viennent alimenter les réflexions autour du plaisir. Chez les oscines, on considère que le chant qui est l’apanage des mâles chez de nombreuses espèces, est impliqué dans la sélection sexuelle. Ainsi, suivant l’espèce considérée, le chant servirait à défendre un territoire et/ou à attirer des partenaires sexuelles potentielles.
Chez les femelles de bruant à gorge blanche, on a observé l’activation des circuits de récompense dans le cerveau des femelles lors de l’écoute des chants. L’activation de ces voies dopaminergiques est souvent associée au plaisir à partir des travaux réalisés chez l’humain. Plus encore, on a montré l’activation de ces circuits cérébraux lors de la production du chant chez les mâles de plusieurs espèces. Jusqu’à présent, cette dimension hédoniste du chant chez les oscines avait été occultée par d’autres aspects plus pragmatiques. Par exemple, il a été proposé que le chant, par sa durée ou sa composition sonore, puisse être un indicateur honnête des capacités reproductives ou cognitives de l’émetteur. Mais une analyse approfondie des travaux menés jusqu’à présent ne permet pas de valider ces hypothèses, en grande partie du fait d’une forte variabilité interspécifique.
Dans certains cas, les vocalisations peuvent aussi avoir une fonction référentielle et renseigner sur des éléments de l’environnement, comme la présence de prédateurs. Chez la mésange japonaise, les oiseaux produisent des cris différents pour désigner différents types de prédateurs.
Chez le toui à lunettes (psittacidé), les individus utilisent des cris distincts pour interpeller chaque individu du groupe. De nombreuses vocalisations ont aussi une fonction phatique dans le maintien du contact, en particulier chez les espèces sociales. Enfin, d’autres fonctions que l’on retrouve chez l’humain posent question chez les animaux non humains et chez les oiseaux tout particulièrement : la fonction conative et la fonction poétique.
Une source d’inspiration
Concernant la fonction conative, c’est la question de l’intentionnalité des vocalisations chez les animaux non humains qui fait débat. Dans les exemples cités ci-dessus (cris de contact chez le diamant mandarin, cris d’alarme de la mésange japonaise), on pourrait interpréter ces cris comme une représentation de l’état interne de l’émetteur, sans volonté de sa part de modifier le comportement du ou des réceveur(s) potentiel(s) et par là même sans avoir forcément besoin de leur prêter des capacités cognitives, comme des attentes ou des intentions.
Pendant des années, l’intentionnalité était considérée comme une caractéristique intrinsèquement liée au langage chez l’humain. Ces dernières années, la liste de critères à remplir pour pouvoir parler d’intentionnalité dans le domaine des communications animales a été revue afin de permettre d’explorer ses aspects phylogénétiques.
Concernant la fonction poétique qui renvoie aussi à la notion d’esthétique, c’est le chant des oscines que l’on peut convoquer pour l’illustrer. Le chant de différentes espèces a inspiré de nombreux compositeurs célèbres comme Ludwig Van Beethoven ou Olivier Messiaen : on retrouve de la musicalité dans certaines de ces productions sonores avec des moments de tension et de relaxation. Le chant pourrait donc être aussi perçu comme une activité artistique, renvoyant à la dimension hédoniste de ce signal évoquée précédemment.
Si les travaux sur le chant des oscines ont jusqu’à présent mis en exergue les parallèles avec le langage chez l’humain, ils peuvent aussi potentiellement alimenter les réflexions autour de la musique, son évolution voire ses aspects phylogénétiques.
Avec leurs vocalisations diverses et variées, les oiseaux occupent une place importante dans l’univers sonore de la planète et nous avons encore beaucoup à découvrir sur ce qu’ils nous en disent.
Retrouvez Sébastien Derégnaucourt pour sa conférence à la Cité des sciences sur le thème de la communication chez les oiseaux, le 16 janvier 2020 à 19 heures.
[ad_2]
Sébastien Derégnaucourt, Éthologiste, professeur des universités, directeur du laboratoire « Éthologie, cognition, développement », Université Paris Nanterre – Université Paris Lumières
Cet article est republié à partir de The Conversation sous licence Creative Commons.